La glycolyse aérobie (catabolisme)
La glycolyse aérobie (catabolisme)
Dernière mise à jour : 2021.11.19
La première étape du catabolisme glucidique comprend la décomposition des macromolécules en sucres simples. Cette décomposition a lieu dans l'intestin grêle. Des molécules organiques sont de cette façon converties en éléments prêts à l'emploi pour l'organisme. Les petites molécules peuvent traverser la paroi intestinale vers la circulation sanguine et finalement entrer dans la cellule.
-
-
les hydrates de carbone d'origine alimentaire sont initialement transformés en glucides simples (glucose) et ensuite absorbés dans la circulation sanguine.
-
ce passage entraîne une élévation de la glycémie, qui stimule une libération insulinique.
-
l'insuline aide l'absorption par le foie et les tissus (= cellules) du glucose circulant.
-
finalement, la glycémie diminue à nouveau et l'insuline en excès est dégradée au niveau hépatique.
-
La dégradation de glucose en dioxyde de carbone (CO2) et eau représente le métabolisme glucidique ou des hydrates de carbone. Ce processus comprend 4 parties : la glycolyse (+ la décarboxylation oxydative), le cycle de l'acide citrique, la chaîne respiratoire et la phosphorylation oxydative.
-
-
45% de tous les sucres absorbés d'origine alimentaire servent au le maintien de la fonction cérébrale, seuls 10% vont vers les muscles. Le reste est impliqué dans le maintien de tous les processus biochimiques.
-
-
-
20% de l'apport oxygénique total dans l'organisme est utilisé dans la génération d'énergie.
-
Sommaire :
 La glycolyse aérobie et anaérobie
La glycolyse aérobie et anaérobie
Aperçu des autres voies métaboliques
La décarboxylation oxydative
Le cycle de l'acide citrique
La chaîne respiratoire
La phosphorylation oxydative
Côté pratique
Schémas
Contenu :
La glycolyse aérobie et anaérobie :
-
-
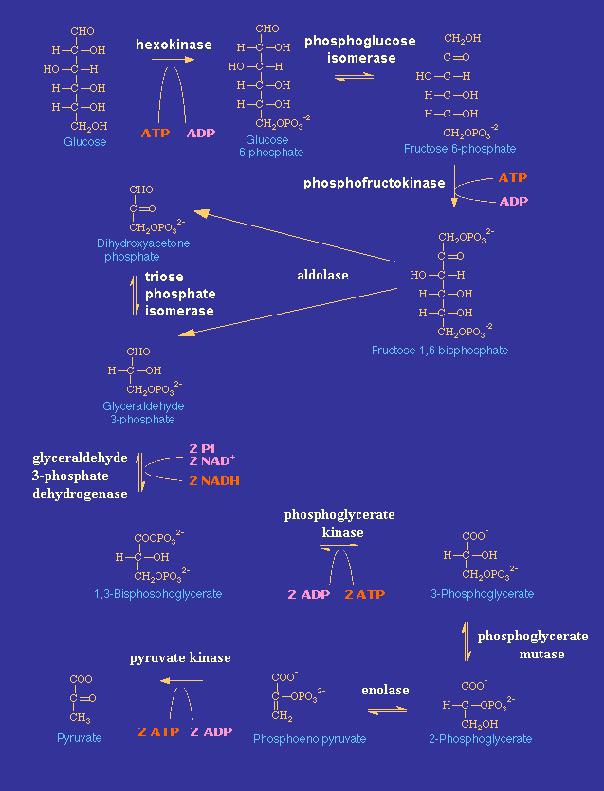
La glycolyse débute dans le cytoplasme. Ce processus dégrade une molécule de glucose en 2 molécules de pyruvate.
-
C6H12O6 + 2 ADP + 2 Pi + 2 NAD+ ---> 2 C3H4O3 + 2 ATP + 2 NADH + 2 H+ + 2 H2O
ou
1 x glucose ----> 2 x pyruvate
La molécule de glucose est d'abord transformée en 2 trioses phosphate avec consommation d'énergie (ATP).
-
-
-
glucose ---> via la hexokinase + ATP ---> glucose 6-phosphate ---> fructose 6-phosphate
-
-
-
-
-
-
fructose 6-phosphate ---> via la phosphofructokinase (PFK) + ATP ---> fructose 1.6-phosphate ---> cission en 2 x glyceraldéhyde 3-P
-
-
-
-
-
-
-
-
les 2 molécules du Glycéraldéhyde 3-phosphate se convertissent en 2 molécules de pyruvate.
-
-
-
-
![]() L'énergie investie sera plus tard récupérée lors de la conversion oxydative du triose-phosphate vers la 3 phosphoglycérate, cédant un proton H+ au coenzyme NAD+ pour former le NADH, la forme réduite.
L'énergie investie sera plus tard récupérée lors de la conversion oxydative du triose-phosphate vers la 3 phosphoglycérate, cédant un proton H+ au coenzyme NAD+ pour former le NADH, la forme réduite.
NAD+ + H+ + 2e- ---> NADH
Lors de la transformation de la phosphoglycérate en pyruvate, le résultat énergétique (ATP) du processus devient positif. Pour chaque molécule de glucose, 2 molécules d'ATP sont consommées mais 4 molécules sont produites. Le résultat est donc positif : un bénéfice de 2 molécules d'ATP!
-
-
Suivant les besoins de l'organisme, le pyruvate est ensuite converti :
-
-
-
-
en CO2, eau et ATP (glycolyse en aérobiose via le cycle de l'acide citrique dans les mitochondries : = décarboxylation via le complexe enzymatique de la pyruvate déshydrogénase en CO2, NADH et en acétylCoA ) : PROCESSUS ASSEZ LENT.
-
-
OU
-
-
-
en acide lactique et ATP (via la glycolyse en aérobiose dans le cytoplasme) : PROCESSUS RELATIVEMENT RAPIDE.
-
-
Chaque route permet la régénération de NADH en NAD+. Cette régénération du NAD+ permet à la glycolyse de se maintenir en absence d'oxygène.
La vitesse d'exécution des processus de glycolyse aérobie et anaérobie diffère : en effet, l'activité enzymatique est plus faible dans les mitochondries que dans le plasma ; en cas d'apport excessif de glucose, plus de lactate (acidification!) et moins d'ATP par phosphorylation oxydative (voir plus loin) seront produits, entraînant un manque d'énergie et des sensations de fatigue.
---> en outre, la combustion rapide du glucose peut induire une hypoglycémie, responsable d'une activation et d'une libération de
-
-
-
glucocorticoïdes, d'adrénaline et de cortisol par les glandes surrénales
-
glucagon par les cellules alpha pancréatiques
-
-
Cette réaction provoque :
-
-
-
-
un état d'hypoglycémie
-
le développement d'un état d'hypoadrénia (épuisement des glandes surrénales)
-
le glycogène hépatique et musculaire tente de compenser les taux sanguins du glucose, le patient est avide de sucreries, en général sous forme d'alimentation raffinée, ce qui rend la correction de la carence des nutriments très difficile ou même impossible
-
l'excès continuel d'acide lactique est à la base d'une stimulation permanente des glandes surrénales, qui maintient un état de stress chronique, causé par l'acidose tissulaire constante (acidose lactique)
-
-
-
![]() Risque d'un état de stress chronique causé par l'excès continuel d'acide lactique (acidose! voir aussi : "Gluconéogenèse").
Risque d'un état de stress chronique causé par l'excès continuel d'acide lactique (acidose! voir aussi : "Gluconéogenèse").
Attention :
-
-
-
-
-
la production d'acide lactique est également impliquée dans la régulation du pH des tissus lors de la formation de nouveaux tissus, de la croissance, de la régénération et récupération tissulaire et de la libération d'oxygène au niveau des cellules tissulaires (voir aussi : "Effet de Bohr").
-
-
-
-
Note:
Cette production d'ATP par la voie de lactate durant la conversion en anaérobiose du glucose représente bien la seule source énergétique des érythrocytes, puisque les globules rouges ne possèdent pas des mitochondries. Quant au lactate toxique formé, il est éliminé durant la gluconéogenèse.
Aperçu des autres voies métaboliques :
A partir du glucose 6-phosphate de la glycolyse :
-
-
---> via la 5-G6P déshydrogénase ---> ribose 5-phosphate ---> nucléotides (dans la voie des pentoses phosphates : pentose phosphate pathway = PPP)
-
-
-
---> oxydation avec le NAD+ ---> inositol 1 phosphate ---> + phosphatidyl ---> Phosphatidyl-Inositol (PI) (---> Système de cascade des phosphatidyl-inositols)
-
-
-
---> UDP-glucose ---> oxydation via la UDP-glucose déshydrogénase ---> acide glucuronique ---> impliqué dans la détoxication.
-
-
-
---> UDP-glucose ---> UDP-galactose ---> utile dans la synthèse des sphingolipides.
-
-
-
---> fructose 6-phosphate ---> via la D glucosamine-P isomérase, Gln ---> D-glucosamine 6-P ---> glucosamine ---> + protéine ---> protéoglycans.
-
-
-
---> fructose 6-phosphate ---> glucosamine ---> via l'AcCoA/B5 (Krebs) ---> acide N-acétylneuraminique (NANA, acide sialique) ---> CMP-acide sialique ---> glycoprotéines ---> protéoglycanes.
-
-
-
---> fructose 6-phosphate ---> 1.3 diphosphate glycéride ---> 2.3 Diphospho-Glycérate (2.3 DPG) (cette voie est activée sur demande : elle joue un rôle dans l'absorption/libération d'O2 par l'Hb : voir "Effet de Bohr").
-
-
-
---> fructose 6-phosphate ---> DHAP (dihydroxyacétone phosphate) : substrat dans la gluconéogenèse ou via la fructolyse ---> lactate.
-
 L'organisme détermine l'équilibre entre protéoglycans et glycoprotéines.
L'organisme détermine l'équilibre entre protéoglycans et glycoprotéines.
La décarboxylation oxydative :
Durant la glycolyse, le glucose est converti en pyruvate. Avant d'entrer dans le cycle de l'acide citrique, le pyruvate subit une décarboxylation et une oxydation combinée (décarboxylation oxydative). Les deux carbones restants deviennent le groupement acétyle (CH3CO) de l'AcétylCoA. Cette réaction est catalysée par le complexe multi-enzymatique de la PDH (pyruvate déshydrogénase).
Cette décarboxylation oxydative est donc la deuxième étape dans le métabolisme des hydrates de carbone :
Après diffusion à travers la membrane mitochondriale, le pyruvate est, en aérobie, converti par réaction de décarboxylation oxydative irréversible en acide acétique activé. Le coenzyme A joue le rôle de molécule porteuse dans la formation du complexe enzymatique Acétyl-Coenzyme A.
C3H4O3 (pyruvate) - CO2 + CoA-SH ---> - 1 Carbone ---> AcétylCoA (= C2)
(Le terme décarboxylation explique la réaction au cours de laquelle un groupement carboxyle sous forme de CO2 est éliminé d'un acide carbonique (R-COOH))
(l'acide acétique, également le produit actif du vinaigre de cidre de pommes, améliore la sensibilité à l'insuline et réduit la formation de graisse en aidant l'organisme à brûler plus efficacement le glucose et à le stocker dans les graisses après les repas)
Le complexe multi-enzymatique PDH
Le complexe de la Pyruvate Déshydrogénase (PDH) est composé de différentes enzymes et nécessite la présence de 5 coenzymes différents : CoA (---> vit B5), TPP (thiamine pyrophosphate ---> vit B1), acide lipoïque, FAD (---> vit B2) et NAD+(---> vit B3).
Le résultat net de la réaction via le complexe multi-enzymatique PDH :
Pyruvate + CoA + NAD+ ---> CO2 + AcCoA + NADH + H+
-
-
-
La TPP accepte le groupement acétyle du pyruvate, le cède à l'acide lipoïque et ensuite à la CoA-SH.
-
-
-
-
-
L'activité du complexe PDH est régulée par son état de phosphorylation. On sait que le Ca2+ et le Mg2+ activent la PDH phosphatase.
-
-
-
-
-
Dans le tissu adipeux, l'insuline stimule l'activité de la PDH ; dans le muscle cardiaque, ce sont les catécholamines (adrénaline, noradrénaline...) qui augmentent l'activité de la PDH.
-
-
-
-
-
Le tabagisme, le mercure (poisson, amalgame dentaire....), arsenic : bloquent l'enzyme déshydrogénase.
-
-
-
-
-
Le NADH produit sera réoxydé par la chaîne respiratoire et permettra la synthèse de 3 ATP.
-
-
1 x glucose ---> 2 pyruvate ---> 2 acétylCoA ---> 2 x NADH ---> 2 x 3 ATP = 6 ATP
![]() La plus grande partie de l'ATP utilisée par les cellules pour maintenir l'homéostasie, est produite par l'oxydation du pyruvate suivi du cycle de l'acide citrique. La conséquence d'une déficience du complexe PDH est donc un déficit énergétique au niveau du SNC.
La plus grande partie de l'ATP utilisée par les cellules pour maintenir l'homéostasie, est produite par l'oxydation du pyruvate suivi du cycle de l'acide citrique. La conséquence d'une déficience du complexe PDH est donc un déficit énergétique au niveau du SNC.
Attention :
-
-
La conversion du pyruvate ---> acétate est une réaction irréversible! Une fois converti, l'acétate ne plus être utilisé pour synthétiser du pyruvate. Le pyruvate ne peut donc être obtenu que par dégradation de grandes biomolécules.
-
![]() La cellule doit donc décider à ce moment :
La cellule doit donc décider à ce moment :
-
-
-
soit d'utiliser le pyruvate provenant de la glycolyse dans la synthèse d'énergie via le cycle de l'acide citrique,
-
soit d'utiliser le pyruvate dans d'autres réactions biochimiques (glycolyse en anaérobie et ensuite dans la gluconéogenèse, lors de la lipogenèse dans le cycle du citrate-pyruvate...).
-
-
Le complexe de la PDH se trouve à ce carrefour et contrôle le flux du pyruvate vers les différentes voies métaboliques.
-
-
Une déficience en ce complexe enzymatique freine évidemment la production de citrate. Elle provoque donc l'arrêt du cycle de l'acide citrique (Krebs). Et comme la plus grande partie des besoins énergétiques (ATP) cellulaires est fournie par l'oxydation du pyruvate dans le cycle de l'acide citrique, une déficience de la PDH entraînera une pénurie d'énergie.
-
-
-
L'excès du pyruvate, formé lors de cette déficience, est converti en lactate (acide lactique) causant une acidification toxique dans le sang (acidose lactique).
-
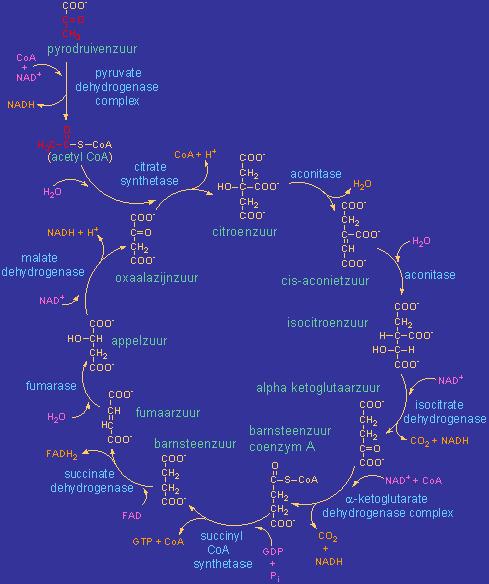
Le cycle de l'acide citrique
(ou cycle de Krebs (découvert par HA. Krebs), ou cycle ATC (des acides tricarboxyliques (3 groupements COOH de l'acide citrique)) :
Le cycle de l'acide citrique est la plaque tournante dans le métabolisme des glucides, des protéines et des graisses.
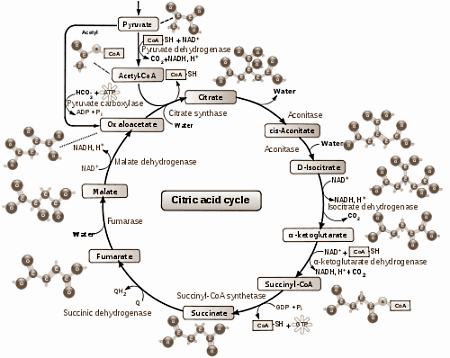
Il se déroule dans la mitochondrie et nécessite la présence d'oxygène (via la respiration : donc l'importance d'une bonne oxygénation tissulaire, d'une bonne circulation pour son apport, et d'un nombre suffisant de globules rouges pour son transport...).
Le cycle de l'acide citrique comprend une série de réactions chimiques qui sont essentielles dans le métabolisme oxydatif du glucose et d'autres glucides simples. Le but du cycle est la formation de NADH et de FADH2 pour la chaîne respiratoire.
Le cycle de l'acide citrique est l'étape finale du catabolisme oxydatif (oxyder = céder des électrons) des carbohydrates, des acides gras et des acides aminés, avec le dioxyde de carbone (CO2) comme produit final du cycle. En réalité, il ne s'agit pas d'une oxydation des molécules, mais d'une réoxydation (régénération) des coenzymes réduits lors de la glycolyse, le NADH et le FADH2, qui est accompagnée d'un transfert d'électrons à l'aide d'une mécanisme porteur, utilisant l'oxygène comme accepteur d'électrons final.
Résultat : Glucide ----> CO2 + H+ et ensuite : NAD+ + H+ + 2e- ---> NADH
-
-
-
chaque cycle produit 1 molécule de GTP (à partir de GDP) (pas d'ATP!). Plus loin, via la chaîne respiratoire et la phosphorylation oxydative, une molécule d'ATP est formée (lors de la libération de 4 électrons).
-
-
Pour chaque molécule de glucose ---> 2 x pyruvate ---> 2 x 1 cycle d'acide citrique ---> 2 x GTP (ATP).
Les produits intermédiaires du cycle servent comme substrats dans la biosynthèse de nouveau matériel cellulaire.
Voir aussi "Cycle de l'acide citrique, les détails" - "Influences externes" - "Influences internes".
La chaîne respiratoire :
Dans la respiration cellulaire normale, les électrons provenant des aliments que nous consommons passent par une série de complexes protéiques dans les mitochondries. Ce processus, connu sous le nom de chaîne de transport d'électrons, aboutit finalement à la production d'ATP, la monnaie énergétique de la cellule. L'oxygène est le dernier accepteur d'électrons dans cette chaîne.
Le transfert des électrons du substrat vers l'oxygène n'est pas direct mais se déroule via une série de réactions, représentée par la chaîne de transport électronique ou la chaîne respiratoire. Elle est localisée dans la membrane interne mitochondriale.
Chaque chaînon de la chaîne réduit le chaînon suivant en s'oxydant lui-même. Chaque étape peut donc subir une réaction d'oxydoréduction réversible. Tel un de système d'escalator. Le couple d'oxydoréduction NAD+/NADH représente le premier chaînon. Un autre couple connu est formé par le FAD/FADH2.
Le flux d'électrons génère également un champ magnétique à l'intérieur des mitochondries. Ce champ magnétique est important pour attirer l'oxygène, qui est essentiel à la production d'énergie, et pour retenir les espèces réactives de l'oxygène dans les mitochondries.
Résultat : 4H+ + O2 + 4e- ---> 2 H2O + énergie pour la conversion ADP + P ---> ATP + Pi
(P : phosphore minéral ; Pi = esters de phosphore)
Ou : C6H12O6 + 6O2 ---> oxydation ---> 6 CO2 + H2O + énergie
Pour chaque molécule de glucose ---> 2 x pyruvate ---> 2 x 1 cycle d'acide citrique :
Cette chaîne de respiration est constituée de 4 complexes enzymatiques :
-
-
I : le complexe de la NADH déshydrogénase (un complexe de flavoprotéines) (vit B3)
-
II : le complexe de la succinate déshydrogénase et de FMN (un complexe de flavoprotéines) (vit B2)
-
III : le complexe de l'ubiquinone déshydrogénase avec les cytochromes b et c1
-
IV : le complexe de la cytochrome oxydase (Fe, Cu, ...)
-
Afin d'optimiser le rôle de la chaîne respiratoire, les éléments suivants sont importants :
-
-
une composition idéale en acides gras essentiels de la membrane mitochondriale
-
un apport suffisant d'alimentation réduite (donc fraîche)
-
une disposition oxygénique suffisante via la respiration cellulaire (bouger!!)
-
une présence suffisante de mévalonate (synthèse de cholestérol) pour la production des cytochromes (donc également un apport suffisant de Cu, Fe...)
-
une synthèse de Q10 suffisante (à partir du cholestérol)
-
apport suffisant de la vitamine B3 (précurseur du NAD)
-
une détoxication : elle permet une accélération de la respiration et une élévation de la phosphorylation oxydative, entraînant une production accrue d'ATP et une stimulation de la synthèse protéique
-
des antioxydants : suite à l'amélioration de la respiration, des fuites d'oxygène peuvent survenir : pathologie RL
-
La phosphorylation oxydative :
Lors du transport d'électrons, les réactions consécutives libèrent de l'énergie : une partie de cette énergie est fixée sous forme d'ATP à partir d'ADP et phosphate : la phosphorylation oxydative.
La phosphorylation oxydative = l'oxydation (= céder des électrons) + phosphorylation (fixation d'un groupement phosphate).
-
-
L'oxydation a lieu durant la réaction réversible suivante :
-
NADH + ubiquinone (CoQ10) <=> NAD+ + ubiquinol en présence de CoQ10 et de Mg2+ .
Normalement la membrane mitochondriale interne est imperméable aux protons. Cependant, au cours du flux d'électrons accompagné du transfert de protons à travers cette membrane interne, il y a formation d'un gradient de protons de part et d'autre de la membrane, ce qui permet la synthèse d'ATP lors d'une réaction catalysée par l'ATP synthase. La chaîne respiratoire et la phosphorylation oxydative sont donc couplées via ce gradient de protons!
-
-
La phosphorylation a lieu durant la réaction réversible suivante :
-
ADP + phosphate ---> ATP synthase ---> ATP + H2O
Les minéraux Cu et Fe sont également impliqués via le complexe des cytochrome oxydases b, c et a (chromoprotéines) dans le transfert des électrons au cours de la chaîne respiratoire.
La phosphorylation oxydative forme la voie de synthèse principale d'ATP, indispensable pour le maintien et la survie cellulaire.
Note:
La présence d'oxygène est cruciale : en cas de carence d'oxygène, le NADH et le FADH ne peuvent pas être réoxydés. Plus de production d'énergie et plus de régénération du NAD+ et du FAD!
-
-
Lorsque les réserves du NAD+ et du FAD sont épuisées, les réactions du cycle de l'acide citrique vont s'arrêter parce que ces coenzymes sont essentiels pour le bon déroulement du cycle! Une pénurie temporaire peut survenir p. ex. lors d'efforts physiques intenses. En cas de manque d'oxygène, le cycle de l'acide citrique fait défaut et le pyruvate s'accumule dans la cellule. Dans cette situation, le pyruvate en excès est transformé en acide lactique, entraînant une acidose sanguine.
-
-
Lorsque les efforts physiques sont réguliers, le tissu musculaire peut s'adapter à un niveau assurant le maintien du cycle de l'acide citrique ainsi que la dégradation rapide d'acide lactique. Dans une telle situation, les poumons et le sang sont capables d'apporter suffisamment d'oxygène, permettant l'oxydation permanente du NADH et FADH.
-
-
-
-
Lorsque le cycle de l'acide citrique est freiné ou bloqué, l'organisme tente de synthétiser son énergie (ATP) via la glycolyse en anaérobiose. Cette voie diminue la glycémie sanguine (hypoglycémie...), stimule les envies de sucreries et active le glucagon (pancréas). L'excès du lactate provoquera une stimulation des glandes surrénales (avec libération d'adrénaline, de cortisol), avec l'éventuelle survenue d'un état de stress chronique (acidose!! voir : "Gluconéogenèse").
-
- La glycolyse «consomme» les sucres de l’alimentation au sein des cellules en produisant une série de métabolites au cours du processus chimique qui décompose le glucose. Les métabolites servent, entre autres, à produire des protéines qui sont nécessaires aux cellules. Un des métabolites, le 1,3-phosphoglycérate cyclique, est un composé très réactif qui peut endommager les protéines et d’autres métabolites. Toutes les cellules humaines ainsi que presque tous les êtres vivants possèdent cette enzyme appelée PARK7. Celle-ci est capable de détruire le dangereux 1,3-phosphoglycérate cyclique. L’inactivation de PARK7 provoque une accumulation de dégâts dans les cellules humaines, mais pas seulement. Les chercheurs ont observé que le même phénomène se produit dans les cellules de souris et de mouches.
Il apparaît que certains cas de maladie de Parkinson seraient dûs à l’inactivation génétique de PARK7. Apparemment, PARK7 peut être facilement inactivé par un stress oxydatif d’origines diverses ![]() .
.
La glycolyse en aérobiose est utilisée en particulier par
les fibres musculaires lentes du type rouge (muscles en métabolisme aérobie) :
-
-
-
source d'énergie : glucose et surtout des acides gras, via la glycolyse aérobie
-
adaptées aux mouvements lents
-
exigent la présence plus élevée de fer (hème) pour les globules rouges (qui sont indispensables dans le transport d'oxygène)
-
consomment du sodium
-
-
Côté pratique :
Le profil sanguin aérobie : normalement le rapport lactate/pyruvate doit être inférieur à 1, le rapport oméga6/oméga3 inférieur à 5, le rapport Fe/Cu environ égal à 1. La présence suffisante de la carnitine est dépendante de celle de la vitamine C et la lysine, la présence des vitamines B12 et B2 est dépendante de celle du facteur intrinsèque gastrique...
Le système aérobie peut être déficient par :
-
-
-
une utilisation insuffisante des muscles en métabolisme aérobie (exercices physiques)
-
une utilisation excessive des muscles en métabolisme anaérobie : étables trop étroites ou de surface insuffisante (en particulier chez des animaux en cage)
-
une déficience en acides gras essentiels : par une mauvaise alimentation
-
un déséquilibre au niveau du métabolisme d'acides gras
-
des déficiences en Cu, Fe, Mn, CoQ10, et vitamines B : des cofacteurs indispensables dans les système aérobie
-
-
Le système aérobie peut être freiné par :
-
-
-
un excès de Cu ... : le Cu déprime le Fe dans les muscles aérobie ; le fructose (fruits...) inhibe l'absorption de Cu
-
la présence de NaF (fluorure de soude) : affaiblit et relâche les muscles aérobie
-
l'acide lactique
-
des antibiotiques
-
-
Schémas :
|
|
La glycolyse
|
|
|
de glucose
à
l'acide pyruvique |
|
|
|
|
Le cycle de l'acide citrique
|
|
|
de l'acide pyruvique
via l'AcétylCoenzyme A
à l'acide citrique
NADH<
avec formation de NADH, FADH2
avec formation de NADH en GTP
|
Le NADH migre vers la chaîne respiratoire (+ O2) et ensuite la conversion d'ADP + Pi ---> ATP (phosphorylation oxydative)
|
le CO2 vers le cycle d'urée > NADH
le CO2 vers le cycle d'urée > NADH
|
|
|
Le transport électronique via le NADH/NAD
|
|
|
|
|
|